

A flagellate-to-amoeboid switch in the closest living relatives of animals
- PMID: 33448265
- PMCID: PMC7895527
- DOI: 10.7554/eLife.61037
A flagellate-to-amoeboid switch in the closest living relatives of animals
Abstract
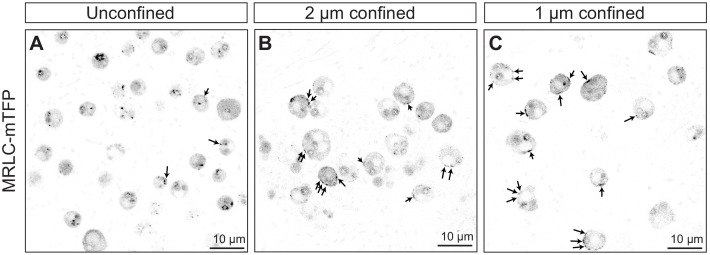
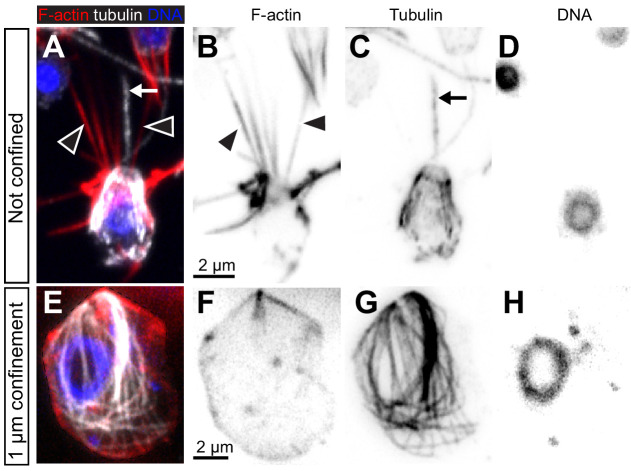
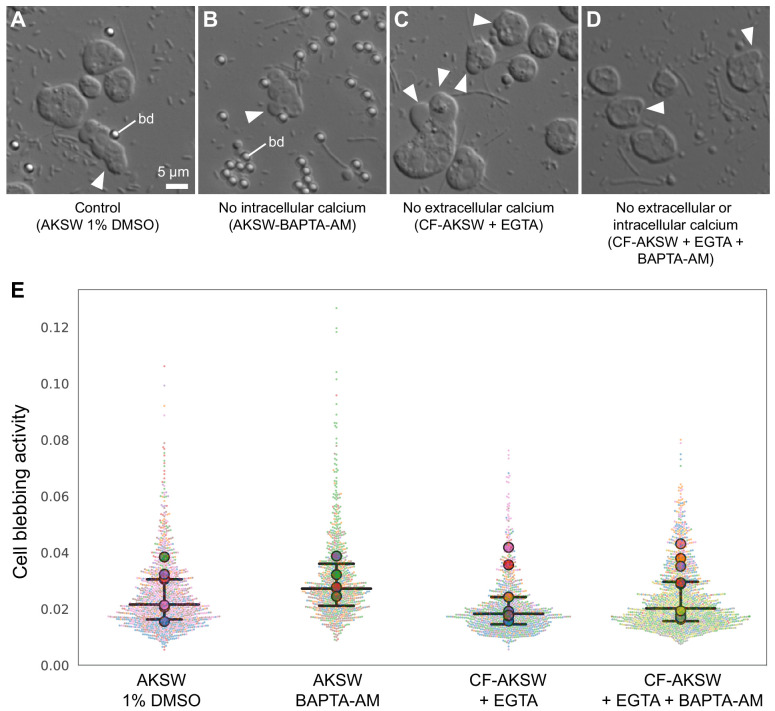
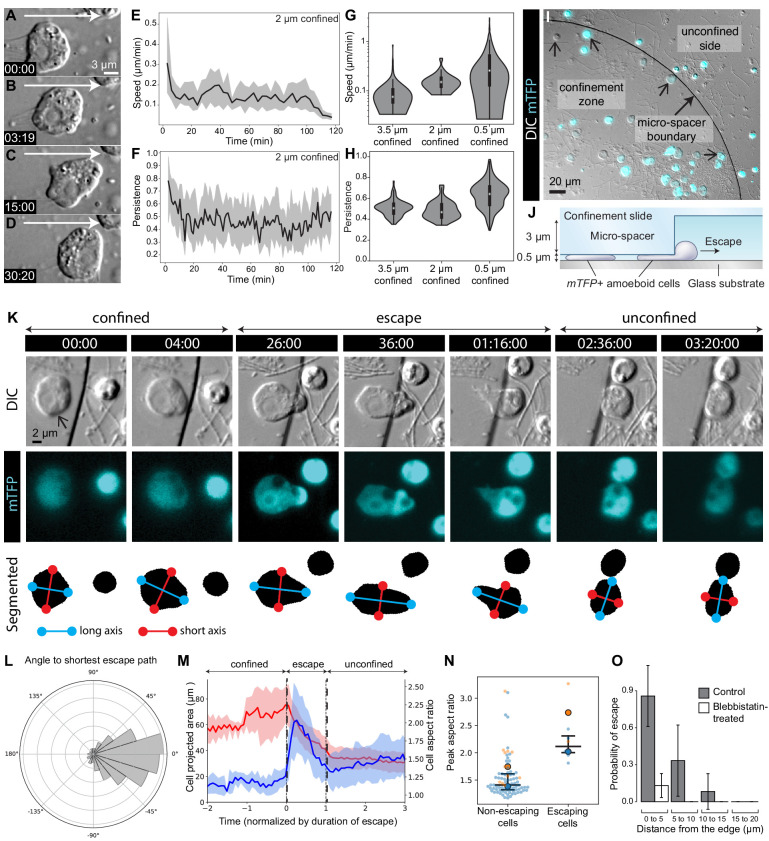
Amoeboid cell types are fundamental to animal biology and broadly distributed across animal diversity, but their evolutionary origin is unclear. The closest living relatives of animals, the choanoflagellates, display a polarized cell architecture (with an apical flagellum encircled by microvilli) that resembles that of epithelial cells and suggests homology, but this architecture differs strikingly from the deformable phenotype of animal amoeboid cells, which instead evoke more distantly related eukaryotes, such as diverse amoebae. Here, we show that choanoflagellates subjected to confinement become amoeboid by retracting their flagella and activating myosin-based motility. This switch allows escape from confinement and is conserved across choanoflagellate diversity. The conservation of the amoeboid cell phenotype across animals and choanoflagellates, together with the conserved role of myosin, is consistent with homology of amoeboid motility in both lineages. We hypothesize that the differentiation between animal epithelial and crawling cells might have evolved from a stress-induced switch between flagellate and amoeboid forms in their single-celled ancestors.
Keywords: Salpingoeca rosetta; animal origins; cell biology; cell type evolution; cellular proprioception; choanoflagellates; evolutionary biology; transdifferentiation.
© 2021, Brunet et al.
Conflict of interest statement
TB, MA, WR, MC, DS, NK No competing interests declared
Figures
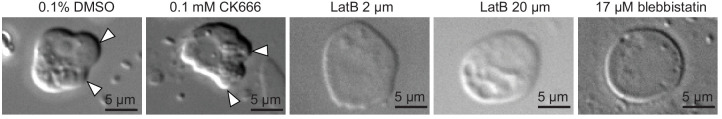
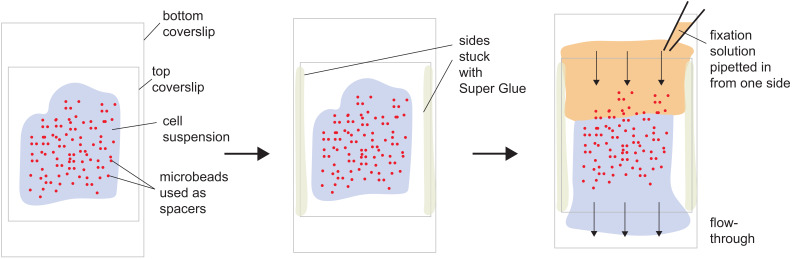
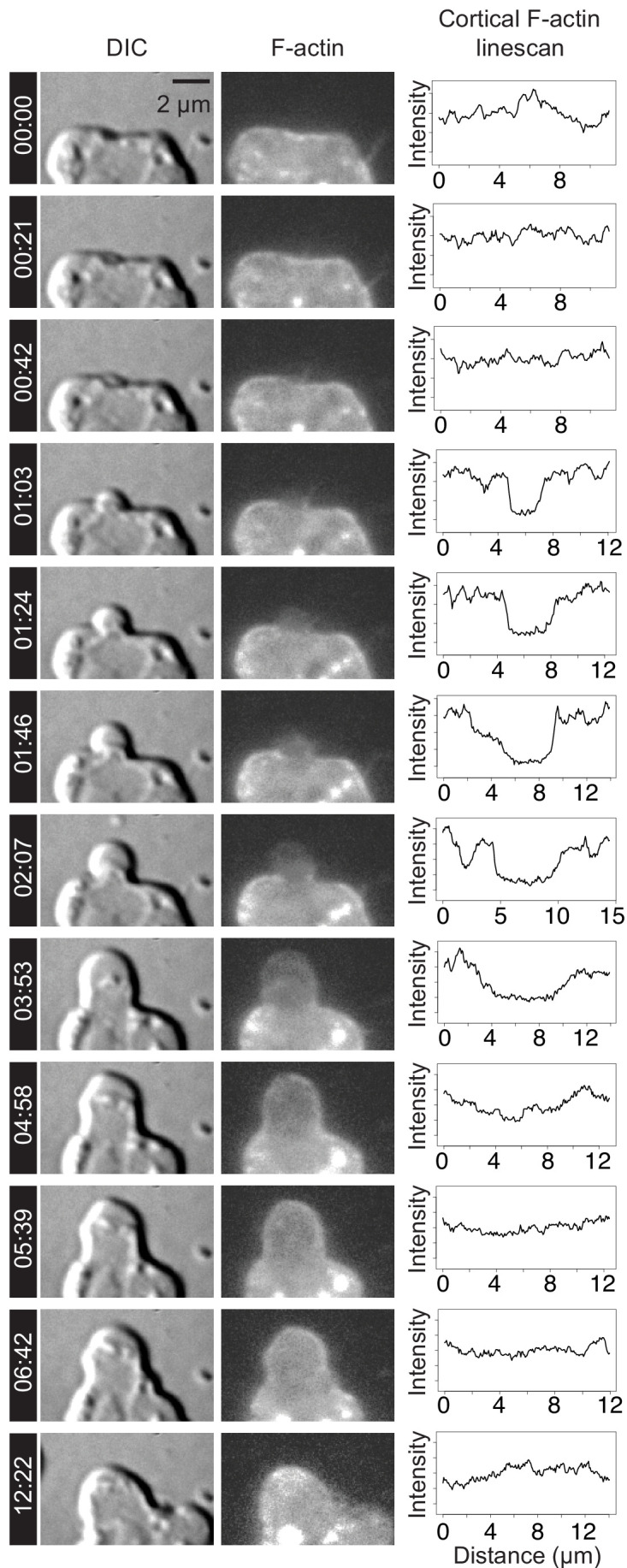
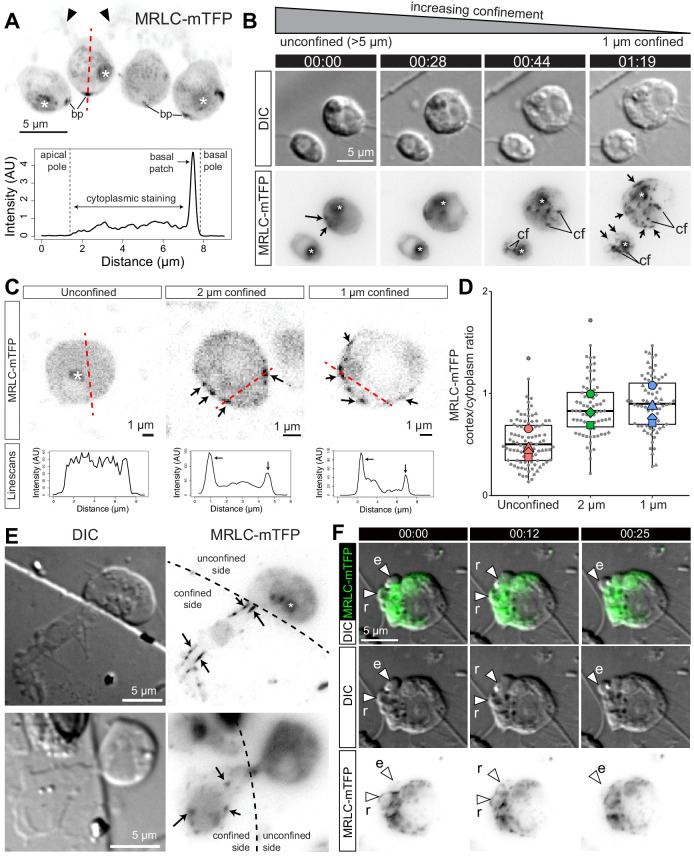
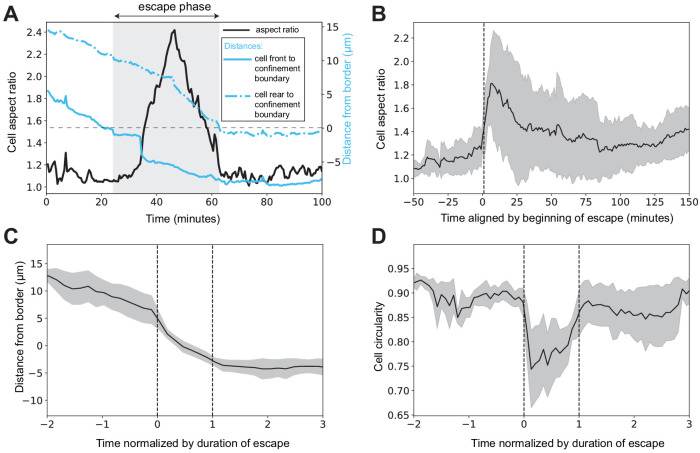
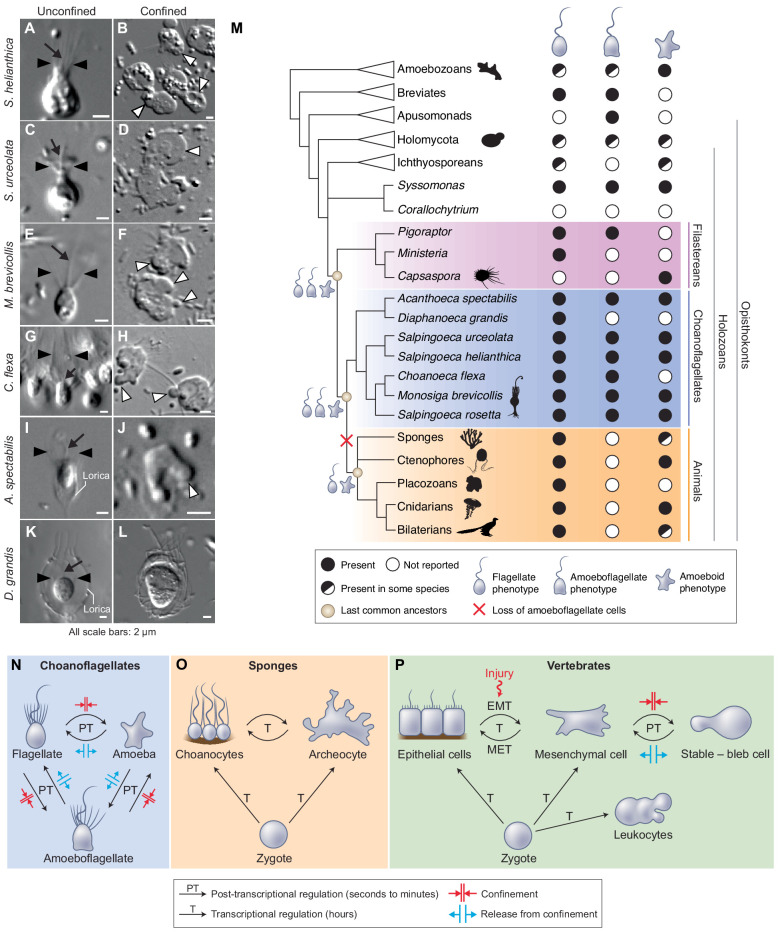
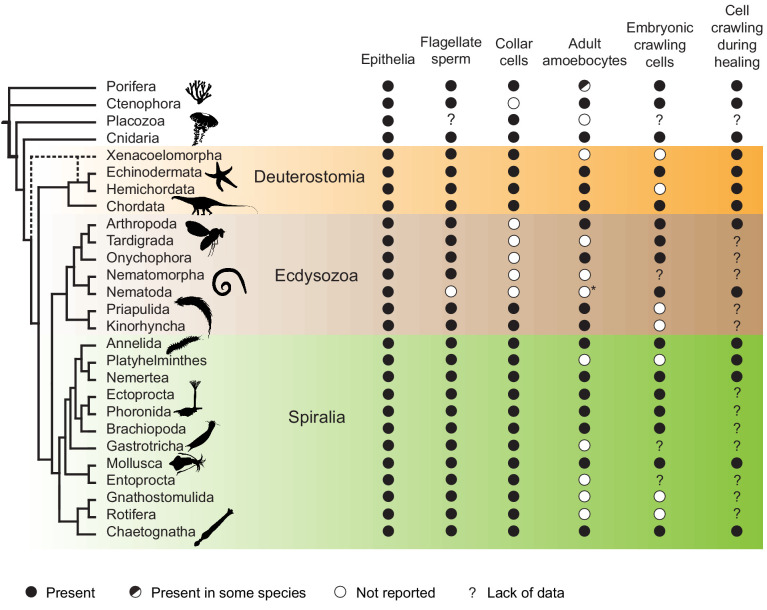
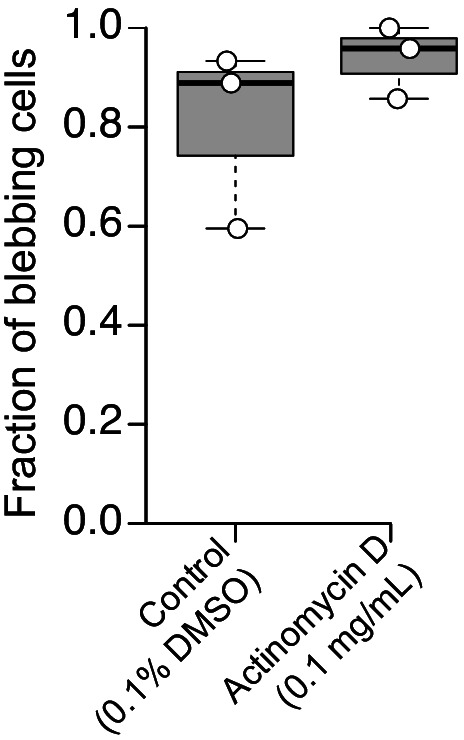
Comment in
-
Evolutionary cell biology: Closest unicellular relatives of animals crawl when squeezed.Curr Biol. 2021 Apr 12;31(7):R353-R355. doi: 10.1016/j.cub.2021.02.012. Curr Biol. 2021. PMID: 33848494
Similar articles
-
Cell differentiation and morphogenesis in the colony-forming choanoflagellate Salpingoeca rosetta.Dev Biol. 2011 Sep 1;357(1):73-82. doi: 10.1016/j.ydbio.2011.06.003. Epub 2011 Jun 12. Dev Biol. 2011. PMID: 21699890 Free PMC article.
-
The architecture of cell differentiation in choanoflagellates and sponge choanocytes.PLoS Biol. 2019 Apr 12;17(4):e3000226. doi: 10.1371/journal.pbio.3000226. eCollection 2019 Apr. PLoS Biol. 2019. PMID: 30978201 Free PMC article.
-
Choanoflagellate models - Monosiga brevicollis and Salpingoeca rosetta.Curr Opin Genet Dev. 2016 Aug;39:42-47. doi: 10.1016/j.gde.2016.05.016. Epub 2016 Jun 17. Curr Opin Genet Dev. 2016. PMID: 27318693 Review.
-
Three-dimensional flagella structures from animals' closest unicellular relatives, the Choanoflagellates.Elife. 2022 Nov 17;11:e78133. doi: 10.7554/eLife.78133. Elife. 2022. PMID: 36384644 Free PMC article.
-
What is it like to be a choanoflagellate? Sensation, processing and behavior in the closest unicellular relatives of animals.Anim Cogn. 2023 Nov;26(6):1767-1782. doi: 10.1007/s10071-023-01776-z. Epub 2023 Apr 17. Anim Cogn. 2023. PMID: 37067637 Free PMC article. Review.
Cited by
-
On the origin of the functional versatility of macrophages.Front Physiol. 2023 Feb 23;14:1128984. doi: 10.3389/fphys.2023.1128984. eCollection 2023. Front Physiol. 2023. PMID: 36909237 Free PMC article.
-
CD44 and Ezrin restrict EGF receptor mobility to generate a novel spatial arrangement of cytoskeletal signaling modules driving bleb-based migration.bioRxiv [Preprint]. 2025 Jan 1:2024.12.31.630838. doi: 10.1101/2024.12.31.630838. bioRxiv. 2025. PMID: 39803565 Free PMC article. Preprint.
-
Placozoan fiber cells: mediators of innate immunity and participants in wound healing.Sci Rep. 2021 Dec 2;11(1):23343. doi: 10.1038/s41598-021-02735-9. Sci Rep. 2021. PMID: 34857844 Free PMC article.
-
Exon Shuffling Played a Decisive Role in the Evolution of the Genetic Toolkit for the Multicellular Body Plan of Metazoa.Genes (Basel). 2021 Mar 8;12(3):382. doi: 10.3390/genes12030382. Genes (Basel). 2021. PMID: 33800339 Free PMC article. Review.
-
Intrinsically disordered proteins and conformational noise: The hypothesis a decade later.iScience. 2023 Jun 15;26(7):107109. doi: 10.1016/j.isci.2023.107109. eCollection 2023 Jul 21. iScience. 2023. PMID: 37408690 Free PMC article. Review.
References
-
- Allan D, Caswell T, Wel CVD. v0.4.2Soft-Matter/Trackpy: Trackpy v0.4.2. 2019 https://zenodo.org/record/3492186#.YAPS9OgzbIU
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
